- 移动端


北京义翘神州科技股份有限公司(Sino Biological Inc.)品牌商
17 年
手机商铺
- NaN
- 0
- 0
- 2
- 2
北京义翘神州科技股份有限公司(Sino Biological Inc.)
入驻年限:17 年
- 联系人:
客服部
- 所在地区:
北京
- 业务范围:
技术服务、试剂、抗体、细胞库 / 细胞培养、ELISA 试剂盒
- 经营模式:
生产厂商
推荐产品




公司新闻/正文
外毒素 vs 内毒素:LPS免疫应答引爆炎症的机制是什么?
293 人阅读发布时间:2026-02-28 11:27
细菌在生长代谢过程中会合成毒素,按其来源、结构、稳定性、毒性和免疫反应可以分为内毒素和外毒素两大类。
1,外毒素
外毒素(Exotoxin)是由活的细菌分泌到细胞外的可溶性蛋白质,主要由革兰氏阳性菌产生。外毒素具有高度性和特异性,是许多致病菌的主要毒力因子。根据其功能可以分为神经毒素、细胞毒素、肠毒素等。外毒素的功能和作用机制见下表:
| 功能 | 作用机制 | 代表毒素 |
| 增强致病性 | 破坏宿主细胞,促进感染 | 破伤风痉挛毒素、白喉毒素 |
| 获取营养 | 释放营养物质 | 溶血素、磷脂酶C |
| 抑制竞争者 | 杀死其他微生物 | 细菌素 |
| 促进传播 | 改变宿主生理(如腹泻) | 霍乱毒素、志贺毒素 |
| 免疫逃逸 | 干扰宿主免疫反应 | 炭疽毒素、葡萄球菌毒素 |
根据外毒素的作用机制和结构特征,还可分为A-B型、膜攻击型和超抗原型。大多数外毒素具有A和B亚单位组成。A亚单位是外毒素的活性部分,决定其毒性。B亚单位是结合亚单位,无毒性但免疫原性强,与宿主表面受体结合,介导A亚单位进入细胞,并发挥其毒性作用。外毒素必须A、B两种亚单位同时存在才有毒性,单独的A不能自行进入宿主细胞,无致病性。
外毒素不稳定,不耐热,一般在60℃加热30分钟可使其灭活。外毒素具有组织特异性,只对特定组织器官造成损害,致死剂量一般在微克或纳克。如1mg肉毒素纯品能杀死2亿只小鼠。
外毒素可刺激宿主产生中和抗体,进而保护宿主免受感染,因此外毒素可作为疫苗原料,经灭活后注射给宿主。外毒素经0.3%甲醛处理后,会成为失去毒性但保留抗原性的类毒素,刺激机体产生外毒素抗体(即抗病毒)。抗毒素可用于治疗和紧急预防。
2,内毒素
内毒素(Endotoxin)主要是细菌细胞壁外膜上的脂多糖(LPS),主要由革兰氏阴性菌死亡或破裂时释放。脂质A是内毒素的活性部分,与宿主细胞表面的受体结合,引发一系列免疫反应。
内毒素稳定,一般需要在180℃以上加热3-4小时或250℃以上加热1-2小时。内毒素通过激活宿主的免疫系统,产生发热、休克、凝血异常等全身性反应。内毒素的致死剂量一般在毫克或微克级别。内毒素不能刺激宿主产生中和抗体,不能作为疫苗的原料,也不能用于免疫诊断。
| 差异维度 | 外毒素 | 内毒素 |
| 来源 | 革兰氏阳性菌为主 | 革兰氏阴性菌 |
| 产生方式 | 活菌分泌 | 细菌死亡后裂解 |
| 化学成分 | 蛋白质 | 脂多糖 |
| 稳定性 | 不稳定,60℃ 30 min | 稳定,180℃ 3-4h |
| 毒性作用 | 强,对组织器官选择性毒害 | 较弱,引起发热、白细胞变化等 |
| 抗原性 | 强,刺激宿主产生抗毒素 | 较弱,不产生中和抗体 |
| 相关疾病 | 霍乱、破伤风、肉毒中毒 | 败血症、脓毒症等 |
3,内毒素诱导炎症反应的分子机制
内毒素是宿主局部及全身性炎症启动过程中的关键分子。其致病作用并非源于直接细胞毒性,而是通过过度激活宿主免疫系统,导致炎症因子过量表达,引发严重的病理损伤。
TLR4介导的经典信号通路
LPS进入血液后,首先与脂多糖结合蛋白(LBP)结合,随后运至髓样细胞表面,与CD14相互作用形成CD14-LPS复合物。该复合物进一步转移至含有髓样分化蛋白MD2的Toll样受体4(TLR4)处,形成CD14-TLR4-MD2复合物。这一过程使免疫细胞特异性识别LPS,进而激活下游信号通路:MyD88和TRIF依赖型通路。
在MyD88依赖型通路中,被激活的MyD88招募白介素-1受体相关激酶(IRAK1、IRAK2、IRAK4)。这些激酶磷酸化后从复合物中解离并与TRAF6结合,通过一系列级联反应激活NF-kB,同时还会激活p38、AP-1、JNK等促炎转录因子的释放。
在TRIF依赖型通路中,TRIF通过招募TRAF3,与TBK1和IKKε形成复合物。该复合物磷酸化后激活IRF3,IRF3与IRF7形成同源或异源二聚体,激活IFN转录。TRIF也可以通过招募TRAF6激活NF-kB。最终促进IL-1β、IL-6、IL-8、MCP-1、TNF-α等炎症细胞因子或趋化因子的大量生成与分泌,驱动炎症反应。
非经典信号通路
LPS还可通过半胱氨酸蛋白酶(Caspase)和瞬时受体电位(TRP)通道激活免疫细胞。在Caspase通路中,LPS激活前体胱天蛋白酶并发生寡聚化,激活Caspase 4/5/11,切割Gasdermin D(GSDMD),破坏细胞膜完整性,促进炎症反应并触发细胞凋亡。同时,Caspase 4/5/11还可能激活NLRPS,促进IL-1β、IL-18分泌。在TRP通道中,TRP通道可快速感知LPS,启动急性炎症反应。
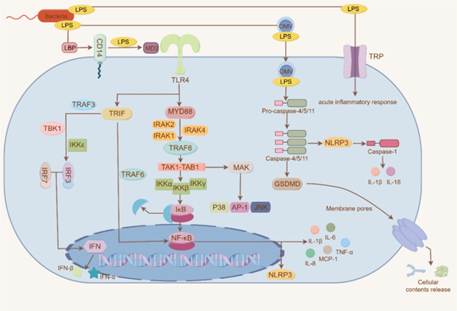
图1. LPS的免疫应答机制
(源自文献:doi:10.1016/j.ijbiomac.2024.135371)
4,内毒素血症
内毒素从感染部位进入血液后,可能形成“内毒素血症”。身体免疫系释放大量炎症因子,产生全身炎症反应。轻者可出现发热、乏力、头痛等不适,严重时可导致器官功能衰竭,甚至威胁生命。内毒素血症是一种因内毒素在血液中大量存在而引起的严重炎症状态,是感染引发败血症和感染性休克的核心机制之一。内毒素本身不会直接杀死细胞,但会引发全身炎症反应综合征(SIRS)、多器官功能障碍综合征(MODS)、血液凝固异常、心源性休克等。
1、致热性
人体对细胞内毒素极为敏感,1-5 ng/kg可导致人体发热。正如上文所说,内毒素引起发热的原因是其作用于白细胞、巨噬细胞,IL-1、IL-6、TNF-α等细胞因子大量表达。促炎因子作为内源性致热源,调节中枢系统的温敏神经元,刺激体温上升。
2、白细胞反应
内毒素进入宿主体内后,会使大量白细胞黏附于毛细血管壁,造成循环血中的白细胞数量急剧下降。随后数小时,内毒素刺激骨髓释放中性粒细胞到血液中,引起白细胞增多,在12-24h达到峰值。
3、弥散性血管内凝血
内毒素促进TNF和IL-1表达,促使白细胞渗出,破坏血凝-抗血凝平衡,导致血栓形成。血小板激活因子PAF引起血小板聚集。总之,内毒素作用于人体后,会导致血小板聚集,触发凝血系统,导致血液凝固,引起弥漫性血管内凝血(DIC)的发生。当DIC严重时,会导致严重的休克症状。
义翘神州ProPure™超低内毒素蛋白及定制服务
义翘神州位于德克萨斯州的生物工程中心(C4B)采用先进的技术和设备生产ProPure™超低内毒素蛋白,内毒素水平低于定量限(BQL)(< 0.05 EU/mg),显著优于行业标准(USP <85>:0.5 EU/mg),适用于临床前动物研究、严格的细胞实验与高敏检测分析。同时,C4B还提供超低内毒素蛋白定制服务,满足临床前和药物开发中对内毒素控制的更高标准需求。
超低内毒素蛋白产品列表(热门靶点)
|
货号 |
分子 |
种属 |
纯度 |
|
10440-H17H-B-UE |
Alkaline Phosphatase |
Human |
≥ 95% (SEC-HPLC) |
|
10279-H08H-UE |
AXL |
Human |
≥ 90% (SEC-HPLC) |
|
91085-C02H-UE |
CD155/PVR |
Cynomolgus |
≥ 95% (SEC-HPLC) |
|
10977-H08H-UE |
CD3e |
Human |
≥ 95% |
|
11150-H08H-UE |
CD69 |
Human |
≥ 90% |
|
CT081-H2508H-UE |
CD79A & CD79B |
Human |
≥ 90% (SEC-HPLC) |
|
11077-H08H-UE |
CEACAM-5/CD66e |
Human |
≥ 95% (SEC-HPLC) |
|
11996-H08H-UE |
c-Kit |
Human |
≥ 90% (SEC-HPLC) |
|
10694-H08H-UE |
EpCAM/TROP1 |
Human |
≥ 90% (SEC-HPLC) |
|
16044-H08H-UE |
FGFR3 |
Human |
≥ 90% (SEC-HPLC) |
|
10004-H08H-UE |
Her2/ERBB2 |
Human |
≥ 90% (SEC-HPLC) |
|
CT022-M08H-UE |
IL12A & IL12B |
Mouse |
≥ 90% |
|
10369-H01H-UE |
IL-13 |
Human |
≥ 90% (SEC-HPLC) |